




















Note: Numbers provided in square brackets in the text refer to the image presented above; image numbers are displayed to the lower left of each image.
General: A perennial forb, 4–10+ dm tall, with 1-3 large compound leaves [1] and a terminal cluster of small white flowers, which develop into a cluster of red or white berries [2–3]. Red baneberry is a shade-tolerant understorey species of moist rich forest types. There are 2 recognized subspecies of Actaea rubra in Canada, but only the typical subsp. rubra occurs in Newfoundland and Labrador. Warning: All portions of this plant are poisonous, but the fruit and rhizomes contain the highest toxic concentration. The leaves and fruit contain a glucoside (ranunculin) and a toxic lactone (protoanemonin) in the form of an irritating yellow oil, while the roots contain a poisonous glucoside (β-sitesterol glucosides). Symptoms of baneberry poisoning from just a few berries include optical migraines, mental confusion, and dizziness, followed by severe abdominal pain, an increased and irregular heartbeat, and general weakness (Bacon 1903). Although poisonous to humans, red baneberry fruits and seeds are eaten by a number of small frugivorous birds and small mammals (Willson 1983).
Key Features: (numbers 1–3 refer to the illustration [4])
1. Leaves 1–3, large, alternate, and ternately compound, with the blade divided into 3 main branches, each of which is pinnately compound; the leaflets of each branch are
lanceolate to ovate, often shallowly lobed, and have coarsely toothed margins.
2. Each flower is composed of 4–10 small sepals and petals, numerous white stamens, and a single superior ovary; individual flowers of the inflorescence are attached by a
slender pedicel.
3. Red or white berries (very rarely pink), 5–11 mm long, ovoid, and somewhat flattened.
Stems: The perennial stem is reduced to a short vertical caudex, about 1 cm in diameter and 6–12 cm long, which persists just below ground level (Mitchell and Dean 1982, Crane 1990). An annual above-ground shoot is produced from the caudex and gives rise to the leaves and inflorescence; the stout petiole-like portion of the stem is about 15–40+ cm long.
Leaves: Cauline, 1–3, alternate, petiolate, and stipulate [5]; the petioles are about 2–24 cm long and the stipules are sheathing. The ternately compound leaf, 2–5 dm long, is divided into 3 main branches [6], each of which is pinnately to bipinnately compound with several lanceolate to broadly ovate leaflets, 2–13 cm long, with attenuate, truncate, or rounded bases, acuminate apices, and shallowly lobed and coarsely serrate margins [7]; image 7 shows detail of the leaflets of one portion of an entire leaf. Leaflets are stalked to nearly sessile, with the lowest pair of leaflets having longer petiolules than upper leaflet pairs; their blades are membranaceous, light to dark green above, paler and usually finely pubescent beneath.
Flowers: Bisexual, with radial symmetry; several (up to 30) white flowers are arranged in a terminal pyramid-shaped (pyramidal) raceme [8–10], 3–5 cm across; the axis and branches of the inflorescence are finely pubescent. The main flowering stalk (peduncle) is 7–15 cm long. Each flower of the raceme is borne on a slender spreading to ascending stalk (pedicel), 0.3–1 cm long and 0.3–0.7 mm in diameter, which is subtended by a small bract. The 4–5 small pale greenish-white sepals are oblanceolate, 2.5–5 mm long, and caducous (falling off as the flower opens); the 4–10 creamy-white petals are small, 2.5–5 mm long, lanceolate to spatulate. The 15–50 stamens have white filaments and anthers; the single pistil is superior with an ovoid white ovary and a sessile 2-lobed stigma, 0.7–1.2 mm in diameter. Pollination is by a variety of small insects (entomophily), especially solitary bees, flies, and small beetles (Pellmyr 1985); the most common pollinator documented from plants in northern Michigan by Pellmyr (1985) was the small European snout beetle, Phyllobius oblongus (L. 1758). Flowers bloom in early summer.
Fruit: An erect cluster, 3–10 cm long, of up to 30 lustrous berries, each 5–11 mm long, ovoid and slightly compressed longitudinally; immature berries are green [11], typically maturing to bright or deep red [12–14] or sometimes white [15–16]. Separate plants of Actaea rubra with red or white fruits are often found growing together [3]. During maturation, some berries appear nearly black [17], due to retention of chlorophyll as the red pigments develop, but when fully mature, these fruits are typically bright or dark red. Plants with white berries have been described as forma neglecta (Gillman) B.L.Rob. The berries are borne on slender green to purplish-black divergent pedicels, up to 2.5 cm long, each subtended by a small lanceolate bract [18]. The fruits also have a small black 'dot' at the end of each berry [18], formed by the persistent stigma, which turns black at maturity. Each fruit contains 10–16 seeds that are hemispherical, flat, and 3–3.6 mm long. The berries have a poisonous white flesh that is bitter to the taste and nausea-inducing (Bacon 1903). While all parts of baneberry plants are poisonous, the berries (and rhizomes) contain higher levels of the poisonous toxins.
Rare plants of Actaea rubra with light pink berries [19] have been found in rural Nova Scotia (K. Walek 2016, pers. comm.) that likely represent a gene mutation, as all of the key traits are consistent with those of Actaea rubra (e.g.: slender green to purplish pedicels, stigma size, and leaf traits) and they have viable seeds).
In Newfoundland and Labrador, red baneberry fruits mature in late July to mid September. Dispersal is by small mammals and frugivorous birds, which are apparently immune to the toxic compounds in the plant; the means of dispersal is through ingestion and defecation (endozoochory). Bird species that have been documented as dispersers, rather than seed predators, include the yellow-bellied sapsucker, American robin, thrushes, and brown thrasher (Crane 1990, Willson 1983), while deer mice, red squirrel, eastern chipmunk, red-backed voles were demonstrated to be primarily predators of baneberry fruits and seeds (Willson 1983).
Ecology and Habitat: Red baneberry occurs with low abundance in moist to wet nutrient-rich forests, mountain maple thickets, and alder swamps in Newfoundland and Labrador. Red baneberry is a shade-tolerant species, but seed germination is most successful in partial shade (Crane 1990). It also occurs in ravines on limestone barrens and in thickets on limestone scree slopes [2-3, 13-14, 16].
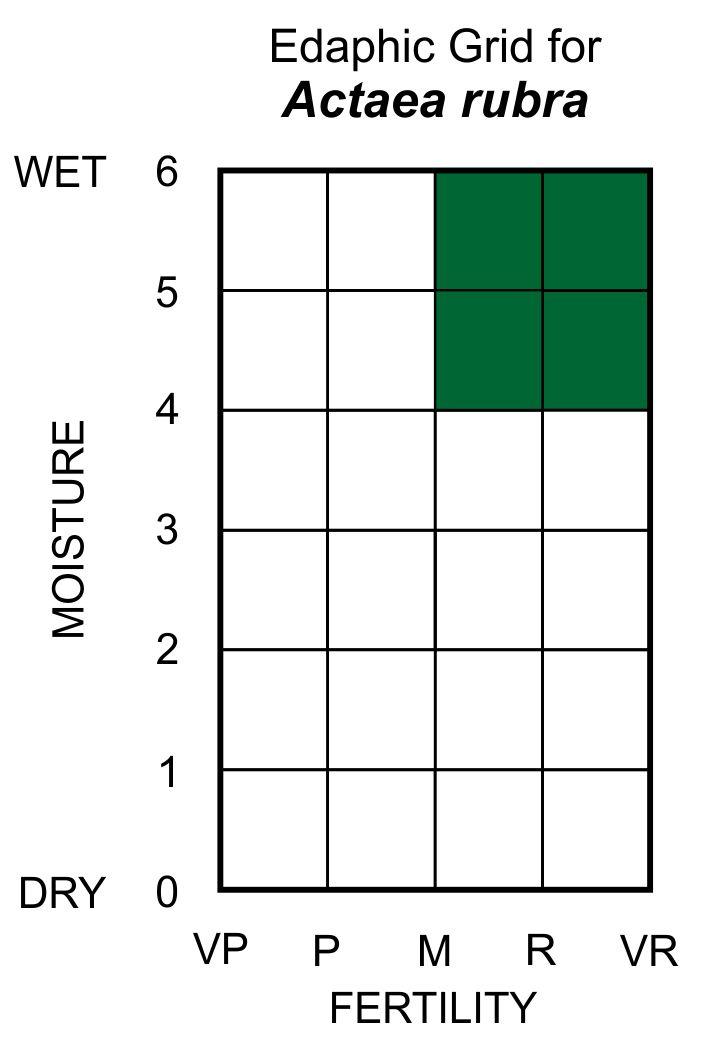
Edaphic Grid: See image [20]: the Edaphic Grid for Actaea rubra.
Forest Types: Red baneberry occurs with low frequency and abundance in the following forest types:
Abietum dryopteretosum (Dryopteris-Balsam Fir Forest Subassociation)
Abietum rubetosum (Rubus-Balsam Fir Forest Subassociation)
Aceretum galietosum (Galium-Mountain Maple Thicket Subassociation)
Alnetum solidagetosum (Solidago-Alder Swamp Subassociation)
Alnetum typicum (Moist Alder Swamp Subassociation)
Succession: Based on studies from elsewhere in North America, red baneberry is known to occur in a wide range of seral stages, after both fire and clear-cutting. Although fires would not be common in its moist habitat, observations indicate that red baneberry is able to persist after fire and disturbance due to its deep-rooted caudex (Crane 1990). Seeds are spread by birds and require nearly 2 years to germinate, with a low 8.8 % germination rate; survival rates of seedlings are somewhat higher in shaded sites (65.3%) than in full sun (50%) (Piper 1986). Hamilton (2006) recorded red baneberry increasing from 33% to 50% presence 10 years after fire in the sub-boreal spruce zone of British Columbia.
Distribution: Actaea rubra is a circumboreal species that occurs in every province of Canada (Brouillet et al. 2010+). The subspecies that occurs across Canada is subsp. rubra, while the range of subs. arguta (Nutt.) Hultén extends from Saskatchewan to British Columbia and the Yukon. Subsp. rubra occurs throughout most of insular Newfoundland, but is lacking from the Avalon Peninsula (Rouleau and Lamoureux 1992); it is most common in rich moist sites in western and northwestern Newfoundland. In Labrador, it extends north to the Churchill River valley and west to Schefferville (Scoggan 1978, Day 1995). In the United States, subsp. rubra occurs across the northern states from Maine to Montana and south to southern Illinois. In the western states, it is replaced by subsp. arguta (USDA, NRCS. 2016).
Similar Species: Leaves of red baneberry resemble those of sweet cicely (Osmorhiza spp.), especially bluntfruit sweet cicely (Osmorhiza depauperata Phil.), which has ternately compound leaves that are membranaceous and finely pubescent. However, the small greenish-white 5-petalled flowers of sweet cicely are arranged in loose umbels and the dry fruit (schizocarps) are narrowly club-shaped (clavate), 1–1.5 cm long, longitudinally ridged, and end in a short blunt tip.
In eastern mainland Canada, a second baneberry species also occurs – doll’s-eyes or white baneberry (Actaea pachypoda Elliott) has white berries, borne on thick pedicels, 0.9–2.2 mm thick, that turn vivid red as the fruits ripen. The common name doll’s-eyes refers to the prominent pupil-like black dot (the persistent stigma), 1.5–2.8 mm across, which marks the apex of each berry. Also, Actaea ×ludovicii B.Boivin, a hybrid of A. rubra and A. pachypoda, has been described from southern Québec; its pedicel thickness and colour are intermediate between the two species (Boivin 1967: 644) and the fruits tend to be sterile (Mitchell and Dean 1982). Plants that are typical of A. pachypoda, but have dark pink to red berries have been described as forma rubrocarpa (Killip) Fernald.